
カイコガと人工身体の融合
昆虫の匂い源定位行動を対象として,昆虫の適応能力を評価するために,昆虫の身体を移動ロボットに置き換えた「昆虫操縦型ロボット」を製作した(図4A)[3, 11].このようなロボットを介した実験系を用いて,カイコガの匂い源定位行動における適応能力を評価した.
この昆虫操縦型ロボットは,行動計測部と行動出力部から構成される.固定されたカイコガがロボット上の発泡スチロール製のボール(直径50 mm)上を歩行し,歩行によって回転するボールの動きをロボットの移動に反映させる仕組みである.行動計測部では,ボールの回転を光学センサにより計測する.行動出力部では,計測したボールの回転から昆虫の移動軌跡を算出し,モータの目標回転量を算出した上でモータを制御し,ロボットの速度制御を行った.カイコガは直進方向で最大速度24.8 mm/s,回転で最大角速度2.1 rad/s 程度となるが,構築した行動出力部は十分にこの性能を満たしていた[3, 11].
風洞内でのフェロモン源定位実験から,定位時間,移動速度ともに,カイコガとロボットでは有意差はなく,このロボットはカイコガの適応行動を調べる上で十分な性能を持っていた[3, 11].
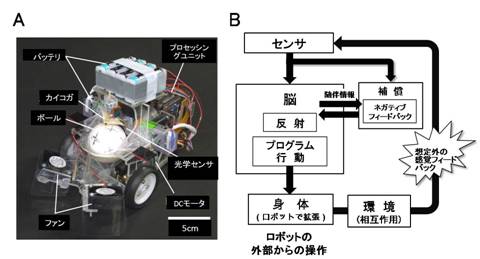
図4.昆虫操縦型ロボット.(A)製作した昆虫操縦型ロボット.(B) ロボットを環境・脳・身体のループに介在させ,ロボットを外部から操作することで,想定外の感覚フィードバックに対して脳がいかに適応的に振る舞うかを評価できる.
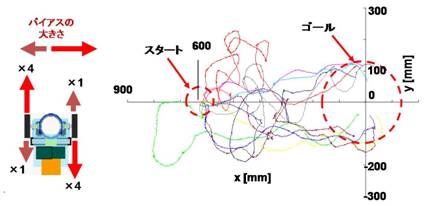
図5.昆虫操縦型ロボットの適応能力.左右非対称に出力を4倍に操作したロボットのフェロモン環境下の移動軌跡.
カイコガの操縦に対して移動ロボットの行動出力を次のように操作してみた.まず,ロボットの左右の両モータの回転速度を通常の0.5倍,2 倍に変更し,フェロモン源定位実験を行った.すると,ロボットはモータ速度の操作に対して,異なる移動軌跡を示したものの,フェロモン源に定位することができた[3, 11].次に,左右非対称に片側の出力のみを4倍に変更してみた(図5).この状況ではカイコガは直進してもロボットは大きく回転し,カイコガの動きによって生じる視覚フィードバックとは全く異なる視覚情報がフィードバックされることになる.はじめはバイアスのかかった方向に大きく回転するものの,瞬時にバイアスのかかった回転が消失し,フェロモン源に向かって定位を始めた(図5).
この結果は,カイコガの行動戦略が,操作による環境−昆虫の相互作用の変化に対しても,十分な匂い源定位能力を維持できる適応能力を持つことを示す.行動出力の操作に対する適応能力としては,プログラム化された行動自体の持つ環境変化に対する頑強性の他に,視覚などの感覚フィードバックによる調節が挙げられる.さらに, 記憶・学習による適応も考えられる.